Hodnocení | Biopsychologie | Srovnávací | Kognitivní | Vývojové | Jazyk | Individuální rozdíly | Osobnost | Filozofie | Sociální metody
Statistiky | Klinické | Vzdělávací | Průmyslové | Profesionální předměty | Světová psychologie |
Biologické: Behaviorální genetika · Evoluční psychologie · Neuroanatomy · Neurochemie · Neuroendokrinologie · Neurovědy · Psychoneuroimunologie · Fyziologická psychologie · Psychofarmakologie (index, přehled) )
| Mozek: Primární sluchová kůra | ||
|---|---|---|
|
||
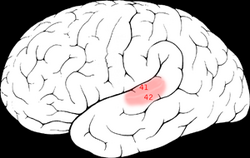
| oblasti Brodmann 41 & 42 lidského mozku. | ||
 |
||
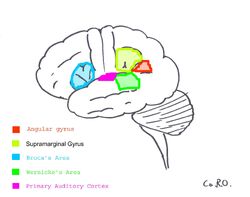
| Primární sluchová kůra je zvýrazněna v purpurová a je známo, že interaguje se všemi oblastmi zvýrazněnými na této neurální mapě. | ||
| latina | “ | |
| Šedá „s | předmět # | |
| Část | ||
| Komponenty | ||
| Tepna | ||
| Žíla | ||
| BrainInfo / UW | ancil-428 | |
| MeSH | ||
Primární sluchová kůra je oblast mozku, která odpovídá za zpracování sluchové (zvukové) informace.
Funkce primární sluchové kůry
Stejně jako u ostatních primárních smyslových kortikálních oblastí dosáhnou sluchové vjemy vnímání pouze tehdy, jsou-li přijaty a zpracovány kortikální oblastí. Důkazy o tom pocházejí ze studií lézí u lidských pacientů, kteří utrpěli poškození kortikálních oblastí nádory nebo mrtvicí, nebo z experimentů na zvířatech, při nichž byly kortikální oblasti deaktivovány chlazením nebo lokálně aplikovanou léčbou. Poškození primární sluchové kůry u lidí vede ke ztrátě jakéhokoli „povědomí“ o zvuku, ale schopnost reflexně reagovat na zvuky zůstává, protože ve sluchovém mozkovém kmeni a středním mozku existuje velká část podkorporálního zpracování.
Neurony ve sluchové kůře jsou organizovány podle frekvence zvuku, na kterou nejlépe reagují. Neurony na jednom konci sluchové kůry nejlépe reagují na nízké frekvence; neurony na druhé straně nejlépe reagují na vysoké frekvence. Existuje několik sluchových oblastí (podobně jako více oblastí ve zrakové kůře), které lze anatomicky odlišit a na základě toho, že obsahují úplnou „frekvenční mapu“. Účel této frekvenční mapy (známý jako tonotopická mapa) není znám a pravděpodobně odráží skutečnost, že senzorický epitel sluchového systému, hlemýžď, je uspořádán podle zvukové frekvence. Sluchová kůra se účastní úkolů, jako je identifikace a segregace sluchových „objektů“ a identifikace umístění zvuku v prostoru.
Skeny lidského mozku ukázaly, že periferní část této oblasti mozku je při pokusu aktivní identifikovat hudební hřiště. Jednotlivé buňky se neustále vzrušují zvuky na konkrétních frekvencích nebo na násobcích této frekvence.
Primární sluchová kůra je přibližně stejná jako Brodmannovy oblasti 41 a 42. Leží v zadní polovině horního spánkového gyrusu a také se ponoří do laterálního sulku jako příčný temporální gyri (také nazývaný Heschl) s gyri).
Primární sluchová kůra se nachází v temporálním laloku. Existují další oblasti lidské mozkové kůry, které se podílejí na zpracování zvuku, v čelních a temenních lalocích. Studie na zvířatech naznačují, že sluchová pole mozkové kůry dostávají vzestupný vstup ze sluchového thalamu a že jsou vzájemně propojeny na stejné a na opačných mozkových hemisférách. Sluchová kůra se skládá z polí, která se navzájem liší strukturou i funkcí.
Počet polí se u různých druhů liší, od 2 u hlodavců až po 15 u opice rhesus. Počet, umístění a organizace polí v lidské sluchové kůře nejsou v tuto chvíli známy. , O čem je známo lidská sluchová kůra pochází ze základů znalostí získaných ze studií na savcích, včetně primátů, používaných k interpretaci elektrofyziologických testů a funkčních zobrazovacích studií mozku u lidí.
Když každý nástroj symfonického orchestru nebo jazzová kapela hraje stejnou notu, kvalita každého zvuku je odlišná – ale hudebník vnímá každou notu se stejnou výškou.Neurony sluchové kůry mozku jsou schopné reagovat na výšku tónu. Studie u opic kosmanů ukázaly, že neurony selektivní ke smola se nacházejí v kortikální oblasti poblíž anterolaterální hranice primární sluchové kůry. Toto umístění oblasti selektivní pro hřiště bylo také identifikováno v nedávných funkčních zobrazovacích studiích u lidí.
Sluchová kůra nedostává pouze vstupy z dolních center a ucha, ale také je poskytuje.
Primární sluchová kůra podléhá modulaci mnoha neurotransmitery, včetně norepinefrinu, u nichž bylo prokázáno, že snižují buněčnou excitabilitu ve všech vrstvách spánkové kůry. Norepinefrin snižuje glutamátergní excitační postsynaptické potenciály na AMPA receptorech aktivací alfa-1 adrenergních receptorů.
Brodmann area 41
Tato oblast je také známá jako přední příčná temporální oblast 41 (H) . Jedná se o dělení cytoarchitektonicky definované časové oblasti mozkové kůry, která zaujímá přední příčný temporální gyrus (H) v břehu laterálního sulku na dorzálním povrchu spánkového laloku. Brodmannova oblast 41 je mediálně ohraničena parainsulární oblastí 52 (H) a laterálně zadní příčnou časovou oblastí 42 (H) (Brodmann-1909).
Brodmannova oblast 42
Toto oblast je také známá jako zadní příčná časová oblast 42 (H). Jedná se o dělení cytoarchitektonicky definované časové oblasti mozkové kůry, která se nachází v břehu laterálního sulku na dorzálním povrchu spánkového laloku. Brodmannova oblast 42 je mediálně ohraničena přední příčnou časovou oblastí 41 (H) a laterálně horní časovou oblastí 22 (Brodmann-1909).
Vztah k sluchovému systému
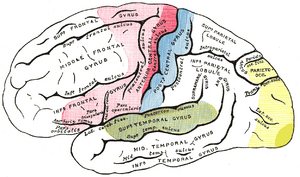
Oblasti lokalizace na bočních povrch polokoule. Oblast motoru v červené barvě. Oblast obecných pocitů modře. Sluchová plocha zeleně. Vizuální oblast žlutá.
Sluchová kůra je nejvíce organizovanou jednotkou zpracování zvuku v mozku. Tato oblast kůry je neurální těžiště sluchu a u lidí jazyk a hudba.
Sluchová kůra je rozdělena do tří samostatných částí, primární, sekundární a terciární sluchové kůry. Tyto struktury jsou tvořeny soustředně kolem sebe, přičemž primární AC je ve středu a terciární AC na vnější straně.
Primární sluchová kůra je tonotopicky uspořádaná, což znamená, že určité buňky ve sluchové kůře jsou citlivé na konkrétní frekvence. Jedná se o fascinující funkci, která se zachovala po většinu konkurzního obvodu. Tato oblast mozku „je považována za identifikaci základních prvků hudby, jako je výška tónu a hlasitost.“ To dává smysl, protože jde o oblast, která přijímá přímý vstup z mediálního geniculárního jádra thalamu. Sekundární sluchová kůra byla indikována při zpracování „harmonických, melodických a rytmických vzorů“. Terciární sluchová kůra údajně integruje vše do celkového zážitku z hudby.
Studie vyvolané odezvy vrozeně neslyšících koťat od Klinke et al. využil místní potenciály pole k měření kortikální plasticity ve sluchové kůře. Tato koťata byla stimulována a měřena proti kontrolní nebo nestimulované vrozeně neslyšící kočce (CDC) a normálním sluchovým kočkám. Potenciály pole měřené pro uměle stimulovaný CDC byly nakonec mnohem silnější než u normální sluchové kočky. To je v souladu se studií Eckarta Altenmullera, kde bylo zjištěno, že studenti, kteří dostali hudební instrukce, měli větší kortikální aktivaci než ti, kteří tak neučinili.
Sluchová kůra vykazuje zvláštní chování týkající se frekvence gama vln. Když jsou subjekty vystaveny třem nebo čtyřem cyklům 40 hertzového kliknutí, objeví se v datech EEG abnormální špička, která není přítomna pro jiné podněty. Špička neuronální aktivity korelující s touto frekvencí není omezena na tonotopickou organizaci sluchové kůry. Předpokládalo se, že se jedná o „rezonanční frekvenci“ určitých oblastí mozku a zdá se, že ovlivňuje také vizuální kůru.
Ukázalo se, že aktivace pásma gama (20 až 40 Hz) je přítomni během vnímání smyslových událostí a procesu rozpoznávání. Kneif a kol. ve své studii z roku 2000 představili předměty s osmi hudebními notami známých melodií, jako jsou Yankee Doodle a Frere Jacques. Šestá a sedmá nota byly náhodně vynechány a elektroencefalogram, stejně jako magnetoencefalogram, byly použity k měření neurálních výsledků. Konkrétně byla přítomnost gama vln vyvolaná sluchovým úkolem měřena z spánků subjektů.Odpověď OSP nebo vynechaná stimulační odpověď byla umístěna v mírně odlišné poloze; 7 mm více vpředu, 13 mm více mediálně a 13 mm lépe oproti kompletním sadám. Záznamy OSP byly také charakteristicky nižší ve vlnách gama, ve srovnání s kompletní hudební sadou. Vyvolávané reakce během šesté a sedmé vynechané noty jsou považovány za představitelné a byly charakteristicky odlišné, zejména na pravé hemisféře. Pravá sluchová kůra se již dlouho ukázala být citlivější na tonalitu, zatímco levá sluchová kůra se ukázala být citlivější na drobné sekvenční rozdíly ve zvuku, konkrétně řeči.
Ukázalo se, že halucinace produkují oscilace, které jsou paralelní (i když ne úplně stejné jako) s frekvenčním rozsahem gama. Sperling ve své studii z roku 2004 ukázal, že sluchové halucinace produkují vlnové délky pásma v rozmezí 12,5-30 Hz. Pásy se vyskytovaly v levé sluchové kůře schizofrenika a byly kontrolovány proti 13 kontrolám (18). To odpovídá studiím lidí, kteří si pamatují píseň v jejich myslích; nevnímají žádný zvuk, ale zažívají melodii, rytmus a celkový zvukový zážitek. Když schizofrenici zažívají halucinace, aktivuje se primární sluchová kůra. To se charakteristicky liší od zapamatování si zvukového stimulu, který jen slabě aktivuje terciální sluchovou kůru. Dedukcí by umělá stimulace primární sluchové kůry měla vyvolat neuvěřitelně skutečnou sluchovou halucinaci. Ukončení celého konkurzu a hudby do terciální sluchové kůry vytváří fascinující spojení zvukových informací. Pokud je tato teorie pravdivá, bylo by zajímavé studovat předmět s poškozeným TAC nebo s uměle potlačenou funkcí. To by bylo velmi obtížné udělat, protože terciární kůra je jednoduše prstenec kolem sekundárního, což je prstenec kolem primárního AC.
Tón je vnímán na více místech než jen ve sluchové kůře; jednou konkrétně fascinující oblastí je rostromediální prefrontální kůra. Janata a kol. Ve své studii z roku 2002 použili stroj fMRI ke studiu oblastí mozku, které byly aktivní během zpracování tonality. Výsledek zobrazil několik oblastí, které se běžně nepovažují za součást procesu konkurzu. Rostromediální prefrontální kůra je podsekcí mediální prefrontální kůry, která vyčnívá do amygdaly a předpokládá se, že pomáhá při potlačení negativních emocí. Mediální prefrontální kůra je považována za základní vývojový rozdíl mezi impulzivním teenagerem a klidným dospělým. Rostromediální prefrontální kůra je citlivá na tonalitu, což znamená, že je aktivována tóny a frekvencemi rezonančních zvuků a hudby. Lze předpokládat, že se jedná o mechanismus, kterým hudba zlepšuje duši (nebo, dává-li se přednost, limbický systém).
Viz také
- Sluchový systém
- Brodmann area
- Účinky hluku na zdraví
- BrainInfo na Washingtonské univerzitě ancil-77: area 41
- BrainInfo na University of Washington ancil-78: area 42
- BrainMaps at UCDavis primary% 20auditory% 20cortex
|
v · d · e
Senzorický systém: sluchové a vestibulární systémy (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vnější ucho |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) • Ušní kanál • Ušní svaly Eardrum (Umbo, Pars flaccida) |
|||||||||||
| středního ucha |
|
|||||||||||
| Vnitřní ucho / (membranózní labyrint, kostnatý labyrint) |
|
|||||||||||
| {| class = „navbox collapsible nowraplinks“ style = „margin: auto;“ | ||||||||||||
|
··
|
||||||||||||
|
||||||||||||
|}
de: Auditiver Cortex]] nl: Auditieve cortex]]
Tato stránka používá licencovaný obsah Creative Commons z Wikipedie (zobrazit autory).