Oppimistulokset
- Kuvaa, kuinka vesipotentiaali vaikuttaa veden kuljettamiseen kasveissa
Kasvit ovat ilmiömäisiä hydrauliikan insinöörejä. Käyttämällä vain fysiikan peruslakeja ja yksinkertaista potentiaalienergian manipulointia kasvit voivat siirtää vettä 116 metriä korkean puun huipulle (kuva 1a). Kasvit voivat myös käyttää hydrauliikkaa riittävän voiman tuottamiseen kivien ja soljen jalkakäytävien halkaisemiseksi (kuva 1b). Kasvit saavuttavat tämän vesipotentiaalin vuoksi.

Kuva 1. Korkeuden ollessa lähellä 116 metriä, (a ) rannikkometsät (Sequoia sempervirens) ovat maailman korkeimpia puita. Kasvien juuret voivat helposti tuottaa tarpeeksi voimaa (b) soljella ja rikkoa betonikäytäviä, mikä on kodinomistajien ja kaupungin kunnossapitoyksiköiden suureksi valitukseksi. (luotto a: Bernt Rostadin tekemä muutos työhön; ansioluettelo b: työn kuljettajien kouluttaminen kuljettajille Safety, Inc. -yritykselle)
Vesipotentiaali mittaa vedessä olevaa potentiaalista energiaa. Kasvifysiologit eivät ole kiinnostuneita minkään tietyn vesipitoisen järjestelmän energiasta, mutta ovat erittäin kiinnostuneita veden liikkumisesta kahden järjestelmän välillä. Käytännössä vesipotentiaali on siis potentiaalienergian ero tietyn vesinäytteen ja puhtaan veden välillä (ilmanpaineessa ja ympäristön lämpötilassa). Vesipotentiaali on merkitty kreikkalaisella kirjaimella ψ (psi) ja se ilmaistaan painoyksikköinä (paine on energiamuoto), nimeltään megapascalit (MPa). Puhtaan veden (Ψpuhdas H2O) potentiaali määritellään määritelmän mukaiseksi nollaksi (vaikka puhdas vesi sisältää paljon potentiaalista energiaa, tämä energia jätetään huomiotta). Kasvien juuressa, varressa tai lehdessä olevan veden vesipotentiaaliarvot ilmaistaan siis suhteessa Ψpuhtaaseen H2O: han.
Kasviliuosten vesipotentiaaliin vaikuttavat liuenneen aineen pitoisuus, paine, painovoima ja matriisiksi kutsutut tekijät vaikutuksia. Vesipotentiaali voidaan jakaa sen yksittäisiin komponentteihin seuraavan yhtälön avulla:
Ψsystem = Ψtotal = Ψs + Ψp + Ψg + Ψm
missä Ψs, Ψp, Ψg ja Ψm tarkoittavat vastaavasti liuenneen aineen, paineen, painovoiman ja matriisipotentiaalia. ”Järjestelmä” voi viitata maaperän veden (maaperän), juuriveden (juuren), varren (varren), lehtien (lehden) tai ilmakehän veden (ilmakehän) vesipotentiaaliin: kumpi vesijärjestelmä on tarkasteltavana Kun yksittäiset komponentit muuttuvat, ne nostavat tai laskevat järjestelmän kokonaisvesipotentiaalia. Kun näin tapahtuu, vesi siirtyy tasapainottua siirtyäkseen järjestelmästä tai osastosta, jolla on suurempi vesipotentiaali, järjestelmään tai osastoon, jolla on pienempi vesipotentiaali. Tämä tuo näiden kahden järjestelmän (ΔΨ) vesipotentiaalieron takaisin nollaan (ΔΨ = 0). Siksi, jotta vesi siirtyisi kasvien läpi maaperästä ilmaan (prosessia kutsutaan transpiroitumiseksi), Ψmaaperän on oltava > Ψroot > Ψstem > Ψleaf > Ψilmapiiri .
Vesi liikkuu vain vasteena ΔΨ: lle, ei vastauksena yksittäisiin komponentteihin, mutta koska yksittäiset komponentit vaikuttavat kokonaismäärään Järjestelmä, manipuloimalla yksittäisiä komponentteja (erityisesti Ψ: itä), kasvi voi hallita veden liikkumista.
Liuotinpotentiaali
Liuotinpotentiaali (Ψs), jota kutsutaan myös osmoottiseksi potentiaaliksi, on negatiivinen kasvisolu ja nolla tislatussa vedessä. Tyypilliset arvot solusytoplasmalle ovat –0,5 – –1,0 MPa. Liuotetut aineet vähentävät vesipotentiaalia (mikä johtaa negatiiviseen Ψw) kuluttamalla osan vedessä käytettävissä olevasta potentiaalisesta energiasta. Liuotetut molekyylit voivat liukenemaan veteen, koska vesimolekyylit voivat sitoutua niihin vetysidosten kautta; öljyn kaltainen hydrofobinen molekyyli, joka ei voi sitoutua veteen, ei voi mennä liuokseen. Liuenneiden molekyylien ja veden välisten vetysidosten energiaa ei enää ole käytettävissä tehdä työtä järjestelmässä, koska se on sitoutunut sidokseen. Toisin sanoen käytettävissä olevan potentiaalienergian määrä vähenee, kun liuenneita aineita lisätään vesipitoiseen järjestelmään. Siten decreasess laskee liuenneen aineen pitoisuuden kasvaessa. Koska Ψs on yksi järjestelmän orsummasta neljästä komponentista, Ψs: n lasku aiheuttaa Ψsumman pienenemisen. Kasvisolun sisäinen vesipotentiaali on negatiivisempi kuin puhdas vesi sytoplasman suuren liuenneen aineen vuoksi (kuva 2). Tämän vesipotentiaalieron vuoksi vesi siirtyy maaperästä kasvin juurisoluihin osmoosiprosessin kautta. Siksi liuenneen aineen potentiaalia kutsutaan joskus osmoottiseksi potentiaaliksi.
Kasvisolut voivat metabolisesti manipuloida Ψ: itä (ja laajemmin Ψ yhteensä) lisäämällä tai poistamalla liuotemolekyylejä. Siksi kasveilla on määräysvalta Ψsummalla niiden kyvyn kautta vaikuttaa aineenvaihduntaan.
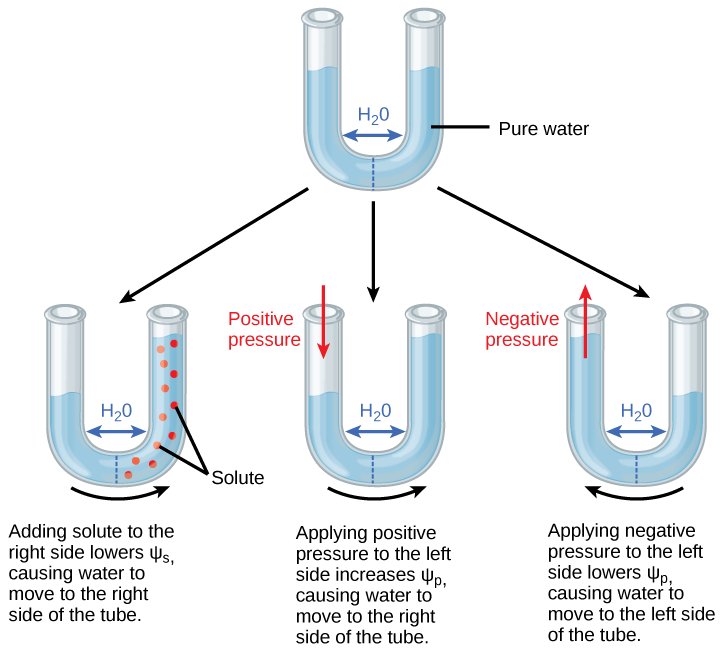
Kuva 2 .Puoliläpäisevä kalvo kahden vesipitoisen järjestelmän välillä
Kuvassa 2 vesi liikkuu alueelta, jolla vesipotentiaali on korkeammalta matalammalle, kunnes tasapaino saavutetaan. Liuenneet aineet (Ψs), paine (Ψp) ja painovoima (Ψg) vaikuttavat kokonaisvesipotentiaaliin putken kummallakin puolella (right yhteensä oikealla tai vasemmalla) ja siten erolla each molemmin puolin (ΔΨ). (Ψm, veden ja kiinteiden substraattien vuorovaikutuksesta johtuva potentiaali jätetään huomiotta tässä esimerkissä, koska lasi ei ole erityisen hydrofiilinen). Vesi liikkuu vastauksena kahden järjestelmän (putken vasen ja oikea puoli) vesipotentiaalin eroon.
Harjoituskysymys
Positiivinen vesipotentiaali sijoitetaan putken vasemmalle puolelle lisäämällä Ψp siten, että vedenpinta nousee oikealla puolella. Voisitko tasata veden tason kummallakin puolella putkea lisäämällä liuenneita aineita, ja jos on, miten?
Painepotentiaali
Painepotentiaali (Ψp), jota kutsutaan myös turgoripotentiaaliksi, voi olla positiivinen tai negatiivinen (kuva 2). Koska paine on energian ilmentymä, sitä korkeampi paine, sitä enemmän potentiaalista energiaa järjestelmässä, ja päinvastoin. Siksi positiivinen Ψp (pakkaus) lisää Ψsummaa ja negatiivinen Ψp (jännitys) pienentää Ψsummaa. Positiivinen paine solujen sisällä on soluseinässä, mikä tuottaa turgoripainetta. Painepotentiaalit ovat tyypillisesti noin 0,6–0,8 MPa, mutta hyvin kastellussa laitoksessa ne voivat olla jopa 1,5 MPa. 1,5 MPa: n Ψp vastaa 210 puntaa neliötuumalle (1,5 MPa x 140 lb-2 MPa-1 = 210 lb / in-2). Vertailun vuoksi useimmat autojen renkaat pidetään 30–34 psi: n paineessa. Esimerkki turgoripaineen vaikutuksesta on lehtien kuihtuminen ja niiden palautuminen kasvien kastelun jälkeen (kuva 3). Vesi häviää lehdistä höyrystymisen kautta (lähestyy Ψp = 0 MPa kuivumiskohdassa) ja palautuu imemällä juuret.
Kasvi voi manipuloida Ψp: tä kykynsä avulla manipuloida Ψ: itä ja prosessin avulla osmoosi. Jos kasvisolu lisää sytoplasmisen liuenneen aineen pitoisuutta, Ψ laskee, Ψ kokonaismäärä pienenee, solun ja ympäröivän kudoksen välinen ΔΨ pienenee, vesi siirtyy soluun osmoosin avulla ja Ψp kasvaa. Ψp on myös epäsuorassa kasvien hallinnassa stomaten avaamisen ja sulkemisen kautta. Stomaaliset aukot antavat veden haihtua lehdestä vähentäen lehden andp- ja Ψsummaa ja lisäämällä ii lehdessä olevan veden ja varren välillä, jolloin vesi virtaa varren lehdestä lehteen.

Kuva 3. Kun (a) kokonaisvesipotentiaali (Ψsumma) on pienempi solujen ulkopuolella kuin sisällä, vesi liikkuu soluista ja kasvi heikkenee. Kun (b) kokonaisvesipotentiaali on kasvisolujen ulkopuolella suurempi kuin sisällä, vesi liikkuu soluihin, mikä johtaa turgoripaineeseen (Ψp) ja pitää kasvin pystyssä. (luotto: muokkaus Victor M. Vicente Selvasin työstä)
Painovoimapotentiaali
Painovoimapotentiaali (Ψg) on aina negatiivinen nollaan laitoksessa, jossa ei ole korkeutta. Se poistaa tai kuluttaa aina potentiaalista energiaa järjestelmästä. Painovoima vetää vettä alaspäin maaperään, mikä vähentää potentiaalisen energian määrää laitoksen vedessä (Ψ yhteensä). Mitä korkeampi kasvi, sitä korkeampi vesipatsas, ja sitä vaikuttavammaksi Ψg tulee. Soluvaikutteisesti ja lyhyissä kasveissa tämä vaikutus on vähäinen ja sivuutetaan helposti. Kuitenkin korkeiden puiden korkeuden, kuten jättimäisen rannikkometsän, painovoima –0,1 MPa m-1 vastaa ylimääräistä 1 MPa: n vastusta, joka on voitettava, jotta vesi pääsee korkeimpien puiden lehtiin. Kasvit eivät pysty manipuloimaan Ψg: tä.
Matriisipotentiaali
Matriisipotentiaali (Ψm) on aina negatiivinen nollaan. Kuivassa järjestelmässä se voi olla jopa –2 MPa kuivassa siemenessä ja nolla vedellä kyllästetyssä järjestelmässä. Veden sitoutuminen matriisiin poistaa tai kuluttaa aina potentiaalista energiaa järjestelmästä. Ψm on samanlainen kuin liuenneen potentiaalin, koska siihen liittyy energian sitominen vesipitoiseen järjestelmään muodostamalla vetysidoksia veden ja jonkin muun komponentin välille. Kuitenkin liuenneessa potentiaalissa muut komponentit ovat liukoisia, hydrofiilisiä liuenneita molekyylejä, kun taas um: ssä muut komponentit ovat kasvisoluseinän liukenemattomia, hydrofiilisiä molekyylejä. Jokaisessa kasvisolussa on selluloosasoluseinä ja soluseinissä oleva selluloosa on hydrofiilinen, mikä tuottaa matriisin veden tarttumiseen: tästä syystä nimi matriisipotentiaali. Ψm on erittäin suuri (negatiivinen) kuivissa kudoksissa, kuten siemenissä tai kuivuudesta kärsineissä maaperissä. Se kuitenkin menee nopeasti nollaan, kun siemen vie vettä tai maaperä kosteutuu.Kasvi ei voi manipuloida Ψm: ää, ja se jätetään yleensä huomiotta hyvin kastelluissa juurissa, varret ja lehdet.
Kokeile
Lisää!
Paranna tätä sivuaLisätietoja