Vurdering | Biopsykologi | Sammenlignende | Kognitiv | Utviklingsmessig | Språk | Individuelle forskjeller | Personlighet | Filosofi | Sosial |
Metoder | Statistikk | Klinisk | Pedagogisk | Industriell | Profesjonelle gjenstander | Verdenspsykologi |
Biologisk: Atferdsmessig genetikk · Evolusjonær psykologi · Nevroanatomi · Nevrokjemi · Nevroendokrinologi · Nevrovitenskap · Psykoneuroimmunologi · Fysiologisk psykologi · Psykofarmakologi (indeks, oversikt) )
| Hjerne: Primær auditiv cortex | ||
|---|---|---|
|
||
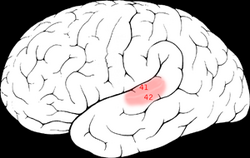
| Brodmann-områder 41 & 42 i den menneskelige hjerne. | ||
 |
||
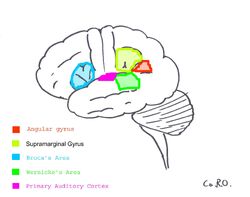
| Primary Auditory Cortex er uthevet i magenta, og har vært kjent for å samhandle med alle områdene som er uthevet på dette nevrale kartet. | ||
| Latin | » | |
| Grå «s | emne # | |
| Del av | ||
| Komponenter | ||
| Arterie | ||
| Vein | ||
| BrainInfo / UW | ancil-428 | |
| MeSH | ||
Den primære auditive cortex er hjernens region som er ansvarlig for behandlingen av auditiv (lyd) informasjon.
Funksjonen til den primære auditive cortex
Som med andre primære sensoriske kortikale områder, oppnår auditive sensasjoner bare oppfatning hvis de mottas og behandles av et kortikalt område. Bevis for dette kommer fra lesjonsstudier hos pasienter hos mennesker som har påført skade på kortikale områder gjennom svulster eller hjerneslag, eller fra dyreforsøk der kortikale områder ble deaktivert ved avkjøling eller medisinsk behandling lokalt. Skade på den primære auditive hjernebarken hos mennesker fører til tap av enhver «bevissthet» om lyd, men en evne til å reagere refleksivt på lyder forblir ettersom det er mye subkortikal prosessering i den auditive hjernestammen og mellomhjernen.
Nevroner i hørebarken er organisert i henhold til lydfrekvensen de reagerer best på. Nevroner i den ene enden av auditiv cortex reagerer best på lave frekvenser; nevroner på den andre reagerer best på høye frekvenser. Det er flere hørselsområder (omtrent som de mange områdene i den visuelle hjernebarken), som kan skilles anatomisk og på bakgrunn av at de inneholder et komplett «frekvenskart.» Hensikten med dette frekvenskartet (kjent som et tonotopisk kart) er ukjent og vil sannsynligvis gjenspeile det faktum at det sensoriske epitelet til det auditive systemet, cochlea, er ordnet i henhold til lydfrekvensen. Den auditive cortex er involvert i oppgaver som å identifisere og adskille auditive «objekter» og identifisere plasseringen av en lyd i rommet.
Menneskelige hjerneskanninger har indikert at en perifer bit av denne hjerneområdet er aktiv når du prøver for å identifisere musikalsk tonehøyde. Individuelle celler blir konsekvent begeistret av lyder ved bestemte frekvenser, eller multipler av den frekvensen.
Den primære auditive cortex er omtrent den samme som Brodmann-områdene 41 og 42. Den ligger i den bakre halvdelen av den overlegne temporal gyrus og dykker også ned i lateral sulcus som den tverrgående temporal gyri (også kalt Heschl » s gyri).
Den primære auditive cortex er lokalisert i den temporale lappen. Det er flere områder av den menneskelige hjernebarken som er involvert i prosessering av lyd, i frontal og parietal lobes. Dyrestudier indikerer at auditive felt i hjernebarken mottar stigende inngang fra den auditive thalamus, og at de er sammenkoblet på samme og på motsatte hjernehalvfinger. Den auditive hjernebarken er sammensatt av felt, som skiller seg fra hverandre i både struktur og funksjon.
Antall felt varierer i forskjellige arter, fra så få som 2 hos gnagere til så mange som 15 i rhesusapen. Antall, plassering og organisering av felt i menneskelig auditiv cortex er foreløpig ikke kjent Hva er kjent om den menneskelige hørselsbarken kommer fra en kunnskapsbase som er innhentet fra studier på pattedyr, inkludert primater, brukt til å tolke elektrofysiologiske tester og funksjonelle avbildningsstudier av hjernen hos mennesker.
Når hvert instrument av symfoniorkesteret eller jazzband spiller samme tone, kvaliteten på hver lyd er forskjellig – men musikeren oppfatter hver tone som den samme tonehøyde.Nevronene i hjernebarken i hjernen er i stand til å svare på tonehøyde. Studier på ape fra marmoset har vist at tonehøyde-selektive nevroner er lokalisert i en kortikal region nær den anterolaterale grensen til primær auditiv cortex. Denne plasseringen av et pitch-selektivt område har også blitt identifisert i nylige funksjonelle avbildningsstudier på mennesker.
Den auditive cortex mottar ikke bare input fra nedre sentre og øret, men gir den også.
Den primære lydbarken er underlagt modulering av en rekke nevrotransmittere, inkludert noradrenalin, som har vist seg å redusere cellulær eksitabilitet i alle lag av timebarken. Noradrenalin reduserer glutamatergiske eksitatoriske postsynaptiske potensialer ved AMPA-reseptorer ved aktivering av alfa-1 adrenerge reseptorer.
Brodmann-område 41
Dette området er også kjent som fremre tverrgående tidsområde 41 (H) . Det er en underavdeling av den cytoarchitecturally-definerte temporale regionen av hjernebarken, som okkuperer den fremre tverrgående temporal gyrus (H) i bredden av lateral sulcus på den dorsale overflaten av den temporale lappen. Brodmann-området 41 avgrenses medialt av parainsularområdet 52 (H) og lateralt av det bakre tverrgående temporale området 42 (H) (Brodmann-1909).
Brodmann-området 42
Dette området er også kjent som bakre tverrgående temporale område 42 (H). Det er en underavdeling av den cytoarchitecturally-definerte temporale regionen av hjernebarken, som ligger i bredden av lateral sulcus på ryggflaten av den temporale lappen. Brodmann-området 42 er avgrenset medialt av det fremre tverrgående tidsområdet 41 (H) og lateralt av det overlegne tidsområdet 22 (Brodmann-1909).
Forholdet til hørselssystemet
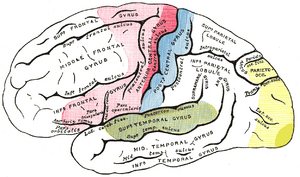
Lokaliseringsområder på siden overflaten av halvkule. Motorområdet i rødt. Område med generelle opplevelser i blått. Hørselsareal i grønt. Visuelt område i gult.
Den auditive cortex er den mest organiserte lydbehandlingsenheten i hjernen. Dette cortexområdet er den nevrale kjernen i hørsel, og hos mennesker, språk og musikk.
Den auditive cortex er delt inn i tre separate deler, den primære, sekundære og tertiære auditive cortex. Disse strukturene er dannet konsentrisk rundt hverandre, med den primære AC i midten og den tertiære AC på utsiden.
Den primære auditive cortex er tonotopisk organisert, noe som betyr at visse celler i auditive cortex er følsomme til bestemte frekvenser. Dette er en fascinerende funksjon som har blitt bevart gjennom det meste av auditionskretsen. Dette området av hjernen «antas å identifisere de grunnleggende elementene i musikk, som tonehøyde og lydstyrke.» Dette er fornuftig, ettersom dette er området som mottar direkte innspill fra thalamus medial geniculate nucleus. Den sekundære auditive cortex har blitt indikert i behandlingen av «harmoniske, melodiske og rytmiske mønstre.» Tertiær auditiv cortex integrerer visstnok alt i den samlede opplevelsen av musikk.
En fremkalt responsstudie av medfødte døve kattunger av Klinke et al. utnyttet lokale feltpotensialer for å måle kortikal plastisitet i auditiv cortex. Disse kattungene ble stimulert og målt mot en kontroll eller un-stimulert medfødt døve katt (CDC) og normale hørselskatter. Feltpotensialene målt for kunstig stimulert CDC var til slutt mye sterkere enn for en normal hørselskatt. Dette er i samsvar med Eckart Altenmüllers studie der det ble observert at studenter som fikk musikalsk instruksjon hadde større kortikalaktivering enn de som ikke gjorde det.
Den auditive cortex viser en merkelig oppførsel knyttet til gammabølgefrekvensen. Når forsøkspersoner utsettes for tre eller fire sykluser med et klikk på 40 hertz, vises en unormal stigning i EEG-dataene, som ikke er tilstede for andre stimuli. Spissen i nevronaktivitet som korrelerer med denne frekvensen er ikke begrenset til den tonotopiske organisasjonen av auditiv cortex. Det er blitt teoretisert at dette er en «resonansfrekvens» for visse områder av hjernen, og ser ut til å påvirke den visuelle cortexen.
Aktivering av gammabånd (20 til 40 Hz) har vist seg å være til stede under oppfatningen av sensoriske hendelser og anerkjennelsesprosessen. Kneif et al. presenterte i sin studie fra 2000 fag med åtte musikalske notater til kjente melodier, som Yankee Doodle og Frere Jacques. Tilfeldigvis ble sjette og syvende tonene utelatt. og et elektroencefalogram, så vel som et magnetoencefalogram ble hver anvendt for å måle nevrale resultater. Spesifikt ble tilstedeværelsen av gammabølger, indusert av den auditive oppgaven ved hånden, målt fra templene til forsøkspersonene.OSP-responsen, eller utelatt stimulusrespons, var lokalisert i en litt annen posisjon; 7 mm mer fremre, 13 mm mer medial og 13 mm mer overlegen i forhold til komplette sett. OSP-opptakene var også karakteristisk lavere i gammabølger, sammenlignet med det komplette musikalske settet. De fremkalte svarene under den sjette og syvende utelatte notatet antas å være forestilt seg, og var karakteristisk forskjellige, spesielt på høyre halvkule. Den høyre auditive cortex har lenge vist seg å være mer følsom for tonalitet, mens den venstre auditive cortex har vist seg å være mer sensitiv for små sekvensielle forskjeller i lyd spesifikt tale.
Hallusinasjoner har vist seg å produsere svingninger som er parallelle (men ikke akkurat det samme som) gammafrekvensområdet. Sperling viste i sin studie fra 2004 at hørselshallusinasjoner produserer båndbølgelengder i området 12,5-30 Hz. Båndene oppstod i venstre auditive cortex av en schizofren og ble kontrollert mot 13 kontroller (18). Dette stemmer overens med studiene av mennesker som husker en sang i deres sinn; de oppfatter ingen lyd, men opplever melodien, rytmen og den generelle opplevelsen av lyd. Når schizofrener opplever hallusinasjoner, er det den primære lydbarken som blir aktiv. Dette er karakteristisk forskjellig fra å huske en lydstimulus, som bare svakt aktiverer tertiær auditiv cortex. Ved deduksjon, bør en kunstig stimulering av den primære auditive cortex fremkalle en utrolig ekte auditiv hallusinasjon. Avslutningen av all audition og musikk i tertiær auditiv cortex skaper en fascinerende sammenheng med lydinformasjon. Hvis denne teorien stemmer, ville det være interessant å studere et emne med en skadet, TAC eller en med kunstig undertrykt funksjon. Dette ville være veldig vanskelig å gjøre da tertiær cortex rett og slett er en ring rundt den sekundære, som er en ring rundt den primære vekselstrømmen.
Tone oppfattes flere steder enn bare hørselsbarken; et spesielt fascinerende område er rostromedial prefrontal cortex. Janata et al., I sin studie fra 2002, brukte en fMRI-maskin for å studere hjernens områder som var aktive under tonalitetsbehandling. Resultatet av dette viste flere områder som normalt ikke anses å være en del av auditionsprosessen. Den rostromediale prefrontale cortexen er en underavdeling av den mediale prefrontale cortexen, som projiserer til amygdalaen, og antas å hjelpe til med å hemme negative følelser. Den mediale prefrontale cortexen antas å være kjernens utviklingsforskjell mellom den impulsive tenåringen og den rolige voksne. Den rostromediale prefrontale cortexen er tonalitetsfølsom, noe som betyr at den aktiveres av tonene og frekvensene av resonante lyder og musikk. Det kan antas at dette er mekanismen som musikk forbedrer sjelen (eller, hvis man foretrekker det limbiske systemet).
Se også
- Auditivt system
- Brodmann-området
- Støyhelseeffekter
- BrainInfo ved University of Washington ancil-77: område 41
- BrainInfo ved University of Washington ancil-78: område 42
- BrainMaps at UCDavis primary% 20auditory% 20cortex
|
v · d · e
Sensorisk system: Auditive og vestibular systems (TA A15.3, GA 10.1029) |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ytre øre |
Pinna (Helix, Antihelix, Tragus, Antitragus, Incisura anterior auris, Earlobe) • Øregang • Aurikulære muskler Trommehinne (Umbo, Pars flaccida) |
|||||||||||
| Mellomøret |
|
|||||||||||
| Indre øre / (membranøs labyrint, benete labyrint) |
|
|||||||||||
| {| class = «navbox sammenleggbare nowraplinks» style = «margin: auto;» | ||||||||||||
|
··
|
||||||||||||
|
||||||||||||
|}
de: Auditiver Cortex]] nl: Auditieve cortex]]
Denne siden bruker Creative Commons lisensiert innhold fra Wikipedia (se forfattere).