Mikrofilamenter
Felles for alle eukaryote celler, disse filamentene er primært strukturelle i funksjon og er en viktig komponent i cytoskelettet, sammen med mikrotubuli og ofte mellomfilamentene. Mikrofilamenter varierer fra 5 til 9 nanometer i diameter og er designet for å bære store mengder spenning. I forbindelse med myosin bidrar mikrofilamenter til å generere kreftene som brukes i cellulær sammentrekning og grunnleggende cellebevegelser. Filamentene gjør det også mulig for en delende celle å klemme seg inn i to celler og er involvert i amoeboidbevegelser av visse typer celler.
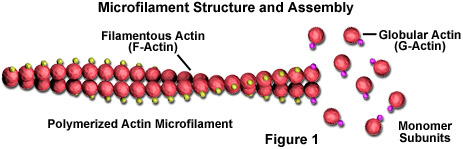
Mikrofilamenter er faste stenger laget av et protein kjent som aktin. Når den først produseres av cellen, vises actin i en kuleform (G-actin; se figur 1). I mikrofilamenter, som også ofte blir referert til som aktinfilamenter, er imidlertid lange polymeriserte kjeder av molekylene flettet sammen i en spiral, og skaper en filamentøs form av proteinet (F-aktin). Alle underenhetene som komponerer et mikrofilament er koblet på en slik måte at de har samme retning. På grunn av dette faktum viser hvert mikrofilament polaritet, de to endene av filamentet er tydelig forskjellige. Denne polariteten påvirker veksthastigheten til mikrofilamenter, den ene enden (betegnet plussenden) monteres og demonteres vanligvis raskere enn den andre (minusenden).
I motsetning til mikrotubuli, som vanligvis strekker seg ut fra sentrum av en celle, blir mikrofilamenter vanligvis kjerneformet ved plasmamembranen. Derfor inneholder periferien (kantene) av en celle vanligvis den høyeste konsentrasjonen av mikrofilamenter. En rekke eksterne faktorer og en gruppe spesielle proteiner påvirker imidlertid mikrofilamentegenskapene og gjør dem i stand til å gjøre raske endringer om nødvendig, selv om filamentene må demonteres helt i en region av cellen og settes sammen et annet sted. Når de finnes rett under plasmamembranen, betraktes mikrofilamenter som en del av cellebarken, som regulerer formen og bevegelsen til celleoverflaten. Derfor spiller mikrofilamenter en nøkkelrolle i utviklingen av forskjellige celleoverflateprojeksjoner (som illustrert i figur 2 ), inkludert filopodia, lamellipodia og stereocilia.
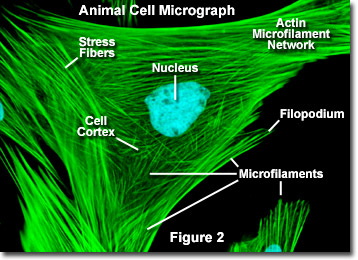
Illustrert i figur 2 er et digitalt fluorescensbilde av en indisk Muntjac-hjortfibroblastcelle farget med fluorescerende prober rettet mot kjernen (blå) og aktin-cytoskeletalt nettverk (grønn). Individuelt er mikrofilamenter relativt fleksible. levende organismer, men aktinfilamentene er vanligvis organisert i større, mye sterkere strukturer av forskjellige tilbehørsproteiner. Den nøyaktige strukturelle formen som en gruppe mikrofilamenter antar, avhenger av deres primære funksjon og de spesielle proteinene som binder dem sammen. For eksempel, i kjernen av overflate fremspring kalt mikrosprikker, er mikrofilamenter organisert i tette parallelle bunter av bunkeproteinet fimbrin. Bunter av filamentene pakkes mindre tett sammen, men når de er bundet av alfa-actinin eller er assosiert med fibroblast-stressfibre (de parallelle grønne fibrene i figur 2). Spesielt mikrofilamentforbindelsene opprettet av noen tverrbindende proteiner resulterer i et nettlignende nettverk eller gelform i stedet for filamentbunter.
I løpet av cellens evolusjonære historie har actin holdt seg relativt uendret. Dette, sammen med det faktum at alle eukaryote celler sterkt avhenger av integriteten til deres aktinfilamenter for å være i stand til å overleve de mange belastningene de står overfor i sitt miljø, gjør aktin til et utmerket mål for organismer som ønsker å skade celler. Følgelig produserer mange planter, som ikke er i stand til å fysisk unngå rovdyr som kanskje vil spise dem eller skade dem på annen måte, giftstoffer som påvirker mobilaktin og mikrofilamenter som en forsvarsmekanisme. Death cap-soppen produserer for eksempel et stoff som kalles falloidin som binder til og stabiliserer aktinfilamenter, noe som kan være dødelig for celler.
TILBAKE TIL DYR CELLESTRUKTUR
TILBAKE TIL PLANT CELLESTRUKTUR
Spørsmål eller kommentarer? Send oss en e-post.
© 1995-2019 av Michael W. Davidson og The Florida State University. Alle rettigheter forbeholdt. Ingen bilder, grafikk, programvare, skript eller applets kan reproduseres eller brukes på noen måte uten tillatelse fra rettighetshaverne. Bruk av dette nettstedet betyr at du godtar alle de juridiske vilkårene som er angitt av eierne.
Dette nettstedet vedlikeholdes av vårt
grafikk & Webprogrammeringsteam
i samarbeid med optisk mikroskopi ved
National High Magnetic Field Laboratory .
Siste modifikasjon: Fredag 13. nov. 2015 kl.13: 18
Antall tilganger siden 1. oktober 2000: 419769
Mikroskoper levert av:
![]()
![]()