Mikrofilament
Gemensamma för alla eukaryota celler, dessa filament har huvudsakligen strukturell funktion och är en viktig komponent i cytoskelettet, tillsammans med mikrotubuli och ofta mellanfilamenten. Mikrofilament sträcker sig från 5 till 9 nanometer i diameter och är utformade för att bära stora mängder spänning. I samband med myosin hjälper mikrofilament att generera de krafter som används i cellulär sammandragning och grundläggande cellrörelser. Filamenten gör det också möjligt för en delande cell att klämma i två celler och är involverade i amoeboidrörelser av vissa typer av celler.
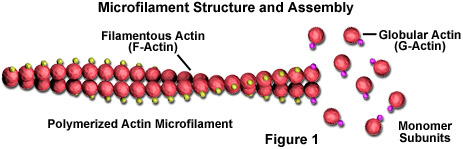
Mikrofilament är fasta stavar tillverkade av ett protein som kallas aktin. När det först produceras av cellen visas aktin i en globform (G-aktin; se figur 1). I mikrofilament, emellertid, som också ofta kallas aktinfilament, är långa polymeriserade kedjor av molekylerna sammanflätade i en spiral, vilket skapar en trådformig form av proteinet (F-aktin). Alla underenheter som komponerar ett mikrofilament är anslutna på ett sådant sätt att de har samma orientering. På grund av detta faktum uppvisar varje mikrofilament polaritet, varvid de två ändarna av filamentet är tydligt olika. Denna polaritet påverkar tillväxthastigheten för mikrofilament, ena änden (benämnd plusänden) monteras och demonteras vanligtvis snabbare än den andra (minusänden).
Till skillnad från mikrotubuli, som vanligtvis sträcker sig ut från centrosomen i en cell, mikrofilament är typiskt kärnformade vid plasmamembranet. Därför innehåller periferin (kanterna) i en cell vanligtvis den högsta koncentrationen av mikrofilament. Ett antal externa faktorer och en grupp speciella proteiner påverkar dock mikrofilamentegenskaperna och gör det möjligt för dem att göra snabba förändringar om det behövs, även om filamenten måste demonteras fullständigt i en region av cellen och återmonteras någon annanstans. När de finns direkt under plasmamembranet betraktas mikrofilament som en del av cellbarken, vilket reglerar formen och rörelsen på cellytan. Följaktligen spelar mikrofilament en nyckelroll i utvecklingen av olika cellytprojektioner (som illustreras i figur 2 ), inklusive filopodia, lamellipodia och stereocilia.
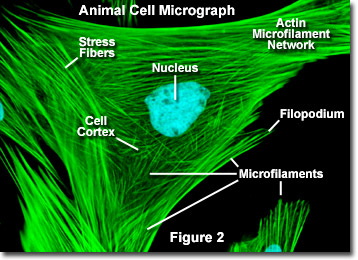
Illustrerad i Figur 2 är en digital fluorescensbild av en indisk Muntjac-hjortfibroblastcell färgad med fluorescerande sonder inriktade på kärnan (blå) och aktin-cytoskeletalt nätverk (grön). Individuellt är mikrofilament relativt flexibla. levande organismer är dock aktinfilamenten vanligtvis organiserade i större, mycket starkare strukturer av olika hjälpproteiner. Den exakta strukturform som en grupp mikrofilament antar beror på deras primära funktion och de specifika proteiner som binder dem ihop. Till exempel, i kärnan av ytutsprång som kallas mikrospikar, organiseras mikrofilament i täta parallella buntar av buntproteinet fimbrin. Buntar av filamenten är mindre tätt packade ihop, men när de är bundna av alfa-aktinin eller är associerade med fibroblast-stressfibrer (de parallella gröna fibrerna i figur 2). I synnerhet resulterar mikrofilamentförbindelserna som skapas av vissa tvärbindande proteiner i ett nätliknande nätverk eller gelform snarare än filamentbuntar.
Under cellens evolutionära historia har aktin förblivit relativt oförändrat. Detta, tillsammans med det faktum att alla eukaryota celler är starkt beroende av integriteten hos deras aktinfilament för att kunna överleva de många påfrestningar de möter i sin miljö, gör aktin till ett utmärkt mål för organismer som försöker skada celler. Följaktligen producerar många växter, som fysiskt inte kan undvika rovdjur som kanske vill äta dem eller skada dem på något annat sätt, toxiner som påverkar cellulärt aktin och mikrofilament som en defensiv mekanism. Dödslocksvampen producerar till exempel ett ämne som kallas falloidin som binder till och stabiliserar aktinfilament, vilket kan vara dödligt för celler.
TILLBAKA TILL Djur CELLSTRUKTUR
TILLBAKA TILL PLANT CELLSTRUKTUR
Frågor eller kommentarer? Skicka oss ett mail.
© 1995-2019 av Michael W. Davidson och Florida State University. Alla rättigheter förbehållna. Inga bilder, grafik, programvara, skript eller applets får reproduceras eller användas på något sätt utan tillstånd från upphovsrättsinnehavarna. Användning av den här webbplatsen innebär att du godkänner alla juridiska villkor som anges av ägarna.
Webbplatsen underhålls av vårt
Graphics & Webbprogrammeringsteam
i samarbete med optisk mikroskopi vid
National High Magnetic Field Laboratory .
Senaste ändring: fredag 13 nov 2015 kl 13:18
Åtkomstantal sedan 1 oktober 2000: 419769
Mikroskop tillhandahållna av:
![]()
![]()