Mikrofilamenter
Fælles for alle eukaryote celler, disse filamenter er primært strukturelle i funktion og er en vigtig komponent i cytoskeletet sammen med mikrotubuli og ofte de mellemliggende filamenter. Mikrofilamenter spænder fra 5 til 9 nanometer i diameter og er designet til at bære store mængder spænding. I forbindelse med myosin hjælper mikrofilamenter med at generere de kræfter, der anvendes i cellulær sammentrækning og grundlæggende cellebevægelser. Filamenterne gør det også muligt for en delende celle at klemme af i to celler og er involveret i amoeboid bevægelser af visse typer celler.
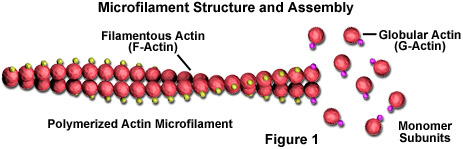
Mikrofilamenter er faste stænger lavet af et protein kendt som actin. Når det først produceres af cellen, vises actin i en kugleform (G-actin; se figur 1). I mikrofilamenter, der imidlertid ofte også omtales som actinfilamenter, er lange polymeriserede kæder af molekylerne sammenflettet i en spiral, hvilket skaber en filamentøs form af proteinet (F-actin). Alle underenheder, der komponerer et mikrofilament, er forbundet på en sådan måde, at de har den samme retning. På grund af dette faktum udviser hvert mikrofilament polaritet, hvor de to ender af filamentet er tydeligt forskellige. Denne polaritet påvirker vækstraten for mikrofilamenter, den ene ende (kaldet plusenden), der typisk samles og skilles ad hurtigere end den anden (minusenden).
I modsætning til mikrotubuli, der typisk strækker sig ud fra centrosomet i en celle, mikrofilamenter kernes typisk ved plasmamembranen. Derfor indeholder periferien (kanterne) af en celle generelt den højeste koncentration af mikrofilamenter. Et antal eksterne faktorer og en gruppe specielle proteiner påvirker imidlertid mikrofilamentkarakteristika og gør dem i stand til at foretage hurtige ændringer, hvis det er nødvendigt, selvom filamenterne skal adskilles helt i en region af cellen og samles et andet sted. Når de findes direkte under plasmamembranen, betragtes mikrofilamenter som en del af cellebarken, som regulerer formen og bevægelsen af celleoverfladen. Derfor spiller mikrofilamenter en nøglerolle i udviklingen af forskellige celleoverfladefremspring (som illustreret i figur 2 ), inklusive filopodia, lamellipodia og stereocilia.
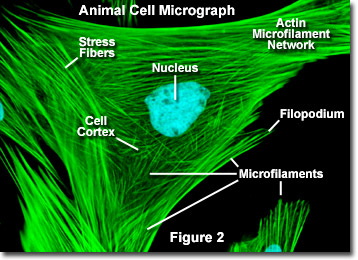
Illustreret i figur 2 er et digitalt fluorescensbillede af en indisk Muntjac-hjortefibroblastcelle farvet med fluorescerende prober målrettet mod kernen (blå) og actin-cytoskeletalt netværk (grøn). Individuelt er mikrofilamenter relativt fleksible. levende organismer organiseres imidlertid actinfilamenterne normalt i større, meget stærkere strukturer af forskellige hjælpeproteiner. Den nøjagtige strukturform, som en gruppe mikrofilamenter antager, afhænger af deres primære funktion og de særlige proteiner, der binder dem sammen. For eksempel i kernen af overfladefremspring kaldet mikrospidser er mikrofilamenter organiseret i tætte parallelle bundter af bundtproteinet fimbrin. Bundter af filamenterne pakkes mindre tæt sammen, men når de er bundet af alfa-actinin eller er forbundet med fibroblast-stressfibre (de parallelle grønne fibre i figur 2). Især resulterer mikrofilamentforbindelserne skabt af nogle tværbindingsproteiner i et weblignende netværk eller gelform snarere end filamentbundter.
I løbet af cellens evolutionære historie har actin forblevet relativt uændret. Dette sammen med det faktum, at alle eukaryote celler stærkt afhænger af integriteten af deres actinfilamenter for at være i stand til at overleve de mange belastninger, de står over for i deres miljø, gør actin til et fremragende mål for organismer, der søger at skade celler. Derfor producerer mange planter, som ikke er i stand til fysisk at undgå rovdyr, der måske vil spise dem eller skade dem på en anden måde, toksiner, der påvirker cellulært actin og mikrofilamenter som en defensiv mekanisme. Death cap-svampen producerer for eksempel et stof kaldet phalloidin, der binder til og stabiliserer actinfilamenter, som kan være fatale for celler.
TILBAGE TIL DYR CELLESTRUKTUR
TILBAGE TIL PLANT CELLESTRUKTUR
Spørgsmål eller kommentarer? Send os en e-mail.
© 1995-2019 af Michael W. Davidson og The Florida State University. Alle rettigheder forbeholdes. Ingen billeder, grafik, software, scripts eller applets må reproduceres eller bruges på nogen måde uden tilladelse fra indehaverne af ophavsretten. Brug af dette websted betyder, at du accepterer alle de juridiske vilkår og betingelser, der er beskrevet af ejerne.
Dette websted vedligeholdes af vores
Grafik & Webprogrammeringsteam
i samarbejde med optisk mikroskopi på
National High Magnetic Field Laboratory .
Sidste ændring: Fredag den 13. november 2015 kl. 13:18
Antal optagelser siden 1. oktober 2000: 419769
Mikroskoper leveret af:
![]()
![]()