Microfilamentos
Común a todas las células eucariotas, estos filamentos tienen una función principalmente estructural y son un componente importante del citoesqueleto, junto con los microtúbulos y, a menudo, los filamentos intermedios. Los microfilamentos varían de 5 a 9 nanómetros de diámetro y están diseñados para soportar grandes cantidades de tensión. En asociación con la miosina, los microfilamentos ayudan a generar las fuerzas utilizadas en la contracción celular y los movimientos celulares básicos. Los filamentos también permiten que una célula en división se pellizque en dos células y participan en los movimientos ameboides de ciertos tipos de células.
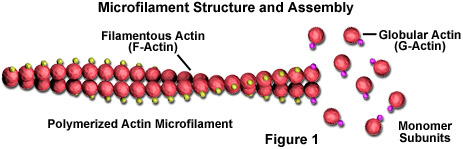
Los microfilamentos son barras sólidas hechas de una proteína conocida como actina. Cuando es producida por primera vez por la célula, la actina aparece en forma globular (actina G; ver Figura 1). Sin embargo, en los microfilamentos, que también se denominan a menudo filamentos de actina, las largas cadenas polimerizadas de las moléculas se entrelazan en una hélice, creando una forma filamentosa de la proteína (F-actina). Todas las subunidades que componen un microfilamento están conectadas de tal manera que tienen la misma orientación. Debido a este hecho, cada microfilamento presenta polaridad, siendo los dos extremos del filamento claramente diferentes. Esta polaridad afecta la tasa de crecimiento de los microfilamentos, un extremo (denominado extremo positivo) generalmente se ensambla y desmonta más rápido que el otro (el extremo negativo).
A diferencia de los microtúbulos, que generalmente se extienden desde el centrosoma de celular, los microfilamentos se nuclean típicamente en la membrana plasmática. Por lo tanto, la periferia (bordes) de una celda generalmente contiene la mayor concentración de microfilamentos. Sin embargo, una serie de factores externos y un grupo de proteínas especiales influyen en las características de los microfilamentos y les permiten realizar cambios rápidos si es necesario, incluso si los filamentos deben desmontarse por completo en una región de la célula y volver a ensamblarse en otra parte. Cuando se encuentran directamente debajo de la membrana plasmática, los microfilamentos se consideran parte de la corteza celular, que regula la forma y el movimiento de la superficie celular. En consecuencia, los microfilamentos desempeñan un papel clave en el desarrollo de diversas proyecciones de la superficie celular (como se ilustra en la Figura 2). ), incluidos filopodios, lamellipodios y estereocilios.
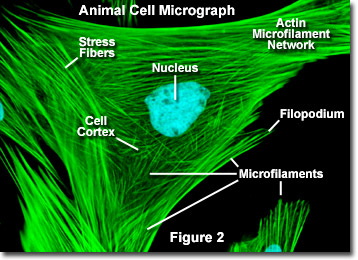
En la Figura 2 se ilustra una imagen digital de fluorescencia de una célula de fibroblastos de piel de ciervo Muntjac indio teñida con sondas fluorescentes dirigidas al núcleo (azul) y la red citoesquelética de actina (verde). Individualmente, los microfilamentos son relativamente flexibles. En las células de En los organismos vivos, sin embargo, los filamentos de actina suelen estar organizados en estructuras más grandes y mucho más fuertes mediante diversas proteínas accesorias La forma estructural exacta que asume un grupo de microfilamentos depende de su función principal y de las proteínas particulares que los unen. Por ejemplo, en el núcleo de las protuberancias de la superficie llamadas microspikes, los microfilamentos se organizan en paquetes paralelos apretados por la proteína fimbrina agrupadora. Sin embargo, los haces de filamentos están menos empaquetados cuando están unidos por alfa-actinina o están asociados con fibras de tensión de fibroblastos (las fibras verdes paralelas en la Figura 2). En particular, las conexiones de microfilamentos creadas por algunas proteínas entrecruzadas dan como resultado una red similar a una red o una forma de gel en lugar de haces de filamentos.
A lo largo de la historia evolutiva de la célula, la actina se ha mantenido relativamente sin cambios. Esto, junto con el hecho de que todas las células eucariotas dependen en gran medida de la integridad de sus filamentos de actina para poder sobrevivir a las muchas tensiones a las que se enfrentan en su entorno, hace que la actina sea un objetivo excelente para los organismos que buscan dañar las células. En consecuencia, muchas plantas, que son incapaces de evitar físicamente a los depredadores que podrían querer comerlas o dañarlas de alguna otra manera, producen toxinas que afectan la actina celular y los microfilamentos como mecanismo de defensa. El hongo del casquete de la muerte, por ejemplo, produce una sustancia llamada faloidina que se une a los filamentos de actina y los estabiliza, que pueden ser fatales para las células.
VOLVER A ANIMAL ESTRUCTURA CELULAR
VOLVER A ESTRUCTURA CELULAR VEGETAL
¿Preguntas o comentarios? Envianos un email.
© 1995-2019 por Michael W. Davidson y la Universidad Estatal de Florida. Reservados todos los derechos. No se pueden reproducir ni utilizar imágenes, gráficos, software, scripts o subprogramas de ninguna manera sin el permiso de los titulares de los derechos de autor. El uso de este sitio web significa que acepta todos los términos y condiciones legales establecidos por los propietarios.
Este sitio web es mantenido por nuestro & Equipo de programación web de
Graphics
en colaboración con Microscopía Óptica en el Laboratorio Nacional de Alto Campo Magnético de
.
Última modificación: viernes 13 de noviembre de 2015 a las 01:18 p.m.
Recuento de accesos desde el 1 de octubre de 2000: 419769
Microscopios proporcionados por:
![]()
![]()