Microfilamenti
Comuni a tutte le cellule eucariotiche, questi filamenti hanno una funzione principalmente strutturale e sono una componente importante del citoscheletro, insieme ai microtubuli e spesso ai filamenti intermedi. I microfilamenti variano da 5 a 9 nanometri di diametro e sono progettati per sopportare grandi quantità di tensione. In associazione con la miosina, i microfilamenti aiutano a generare le forze utilizzate nella contrazione cellulare e nei movimenti cellulari di base. I filamenti consentono inoltre a una cellula in divisione di dividersi in due cellule e sono coinvolti nei movimenti ameboidi di alcuni tipi di cellule.
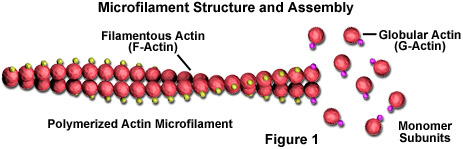
I microfilamenti sono barre solide costituite da una proteina nota come actina. Quando viene prodotta per la prima volta dalla cellula, l’actina appare in una forma globulare (G-actina; vedi Figura 1). Nei microfilamenti, tuttavia, che sono spesso indicati anche come filamenti di actina, lunghe catene polimerizzate delle molecole si intrecciano in un’elica, creando una forma filamentosa della proteina (F-actina). Tutte le subunità che compongono un microfilamento sono collegate in modo tale da avere lo stesso orientamento. A causa di questo fatto, ogni microfilamento mostra polarità, le due estremità del filamento essendo nettamente diverse. Questa polarità influisce sul tasso di crescita dei microfilamenti, un’estremità (denominata estremità positiva) in genere si monta e si smonta più velocemente dell’altra (estremità meno).
A differenza dei microtubuli, che tipicamente si estendono dal centrosoma di un cellula, i microfilamenti sono tipicamente nucleati sulla membrana plasmatica. Pertanto, la periferia (bordi) di una cellula contiene generalmente la più alta concentrazione di microfilamenti. Tuttavia, una serie di fattori esterni e un gruppo di proteine speciali influenzano le caratteristiche dei microfilamenti e consentono loro di apportare modifiche rapide se necessario, anche se i filamenti devono essere completamente smontati in una regione della cellula e riassemblati da qualche altra parte. Quando si trovano direttamente sotto la membrana plasmatica, i microfilamenti sono considerati parte della corteccia cellulare, che regola la forma e il movimento della superficie della cellula. Di conseguenza, i microfilamenti svolgono un ruolo chiave nello sviluppo di varie proiezioni della superficie cellulare (come illustrato nella Figura 2 ), inclusi filopodi, lamellipodi e stereocilia.
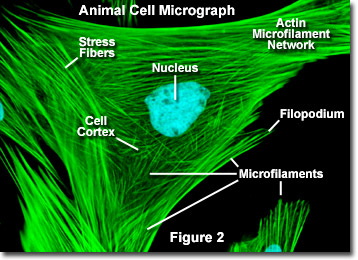
Nella Figura 2 è illustrata un’immagine digitale a fluorescenza di un fibroblasto di pelle di cervo Muntjac indiano colorato con sonde fluorescenti che mirano al nucleo (blu) e alla rete citoscheletrica dell’actina (verde). Individualmente, i microfilamenti sono relativamente flessibili. Nelle cellule di organismi viventi, tuttavia, i filamenti di actina sono solitamente organizzati in strutture più grandi e molto più forti da varie proteine accessorie.La forma strutturale esatta che assume un gruppo di microfilamenti dipende dalla loro funzione primaria e dalle proteine particolari che li legano insieme. Ad esempio, nel nucleo delle sporgenze superficiali chiamate microspike, i microfilamenti sono organizzati in fasci paralleli stretti dalla proteina legante fimbrin. Tuttavia, i fasci dei filamenti sono meno fitti insieme quando sono legati da alfa-actinina o sono associati a fibre di stress dei fibroblasti (le fibre verdi parallele nella Figura 2). In particolare, le connessioni dei microfilamenti create da alcune proteine reticolanti danno come risultato una rete simile a una rete o una forma di gel piuttosto che fasci di filamenti.
Nel corso della storia evolutiva della cellula, l’actina è rimasta relativamente invariata. Questo, insieme al fatto che tutte le cellule eucariotiche dipendono fortemente dall’integrità dei loro filamenti di actina per essere in grado di sopravvivere ai numerosi stress che devono affrontare nel loro ambiente, rende l’actina un ottimo bersaglio per gli organismi che cercano di danneggiare le cellule. Di conseguenza, molte piante, che non sono in grado di evitare fisicamente i predatori che potrebbero volerli mangiare o danneggiarli in qualche altro modo, producono tossine che agiscono sull’actina cellulare e sui microfilamenti come meccanismo difensivo. Il fungo del cappuccio della morte, ad esempio, produce una sostanza chiamata falloidina che si lega e stabilizza i filamenti di actina, che possono essere fatali per le cellule.
TORNA ALL’ANIMALE STRUTTURA CELLULARE
TORNA ALLA STRUTTURA CELLULARE VEGETALE
Domande o commenti? Mandaci un email.
© 1995-2019 di Michael W. Davidson e The Florida State University. Tutti i diritti riservati. Nessuna immagine, grafica, software, script o applet può essere riprodotta o utilizzata in alcun modo senza l’autorizzazione dei titolari del copyright. L’uso di questo sito Web implica l’accettazione di tutti i Termini e condizioni legali stabiliti dai proprietari.
Questo sito Web è gestito dal nostro
Graphics & Team di programmazione Web
in collaborazione con la microscopia ottica presso il Laboratorio nazionale
campo magnetico elevato .
Ultima modifica: venerdì 13 novembre 2015 alle 13:18
Numero di accessi dal 1 ° ottobre 2000: 419769
Microscopi forniti da:
![]()
![]()