Microfilamenten
Gemeenschappelijk voor alle eukaryote cellen, deze filamenten zijn primair structureel in functie en zijn een belangrijk onderdeel van het cytoskelet, samen met microtubuli en vaak de tussenliggende filamenten. Microfilamenten hebben een diameter van 5 tot 9 nanometer en zijn ontworpen om grote hoeveelheden spanning te dragen. In combinatie met myosine helpen microfilamenten om de krachten te genereren die worden gebruikt bij cellulaire contractie en basale celbewegingen. De filamenten zorgen er ook voor dat een deelcel zich in twee cellen kan samenknijpen en zijn betrokken bij amoeboïde bewegingen van bepaalde typen cellen.
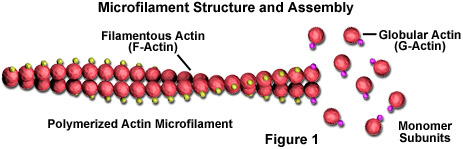
Microfilamenten zijn stevige staafjes gemaakt van een eiwit dat bekend staat als actine. Wanneer het voor het eerst door de cel wordt geproduceerd, verschijnt actine in een bolvorm (G-actine; zie Figuur 1). In microfilamenten, die ook vaak actine-filamenten worden genoemd, zijn lange gepolymeriseerde ketens van de moleculen met elkaar verweven in een helix, waardoor een filamenteuze vorm van het eiwit (F-actine) ontstaat. Alle subeenheden waaruit een microfilament bestaat, zijn zo verbonden dat ze dezelfde oriëntatie hebben. Vanwege dit feit vertoont elk microfilament polariteit, waarbij de twee uiteinden van het filament duidelijk verschillend zijn. Deze polariteit beïnvloedt de groeisnelheid van microfilamenten, waarbij het ene uiteinde (het plus-uiteinde genoemd) doorgaans sneller wordt gemonteerd en gedemonteerd dan het andere (het min-uiteinde).
In tegenstelling tot microtubuli, die zich doorgaans uitstrekken vanaf het centrosoom van een cel, worden microfilamenten typisch genucleëerd op het plasmamembraan. Daarom bevat de periferie (randen) van een cel over het algemeen de hoogste concentratie microfilamenten. Een aantal externe factoren en een groep speciale eiwitten beïnvloeden echter de eigenschappen van microfilamenten en stellen hen in staat om indien nodig snel veranderingen aan te brengen, zelfs als de filamenten in het ene deel van de cel volledig moeten worden gedemonteerd en ergens anders weer in elkaar moeten worden gezet. Wanneer microfilamenten direct onder het plasmamembraan worden aangetroffen, worden ze beschouwd als onderdeel van de celcortex, die de vorm en beweging van het celoppervlak reguleert. Bijgevolg spelen microfilamenten een sleutelrol bij de ontwikkeling van verschillende celoppervlakteprojecties (zoals geïllustreerd in figuur 2 ), inclusief filopodia, lamellipodia en stereocilia.
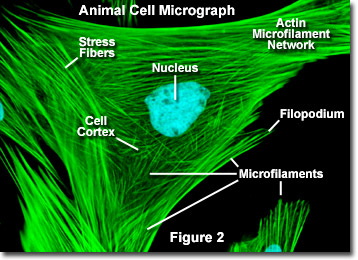
Geïllustreerd in figuur 2 is een digitale fluorescentie-afbeelding van een Indiase Muntjac-fibroblastcel van hertenhuid, gekleurd met fluorescerende sondes die gericht zijn op de kern (blauw) en het actine-cytoskeletnetwerk (groen). Individueel zijn microfilamenten relatief flexibel. In de cellen van levende organismen, de actinefilamenten zijn echter meestal georganiseerd in grotere, veel sterkere structuren door verschillende accessoire-eiwitten De exacte structurele vorm die een groep microfilamenten aanneemt, hangt af van hun primaire functie en de specifieke eiwitten die ze aan elkaar binden. In de kern van oppervlakte-uitsteeksels die microspikes worden genoemd, worden microfilamenten bijvoorbeeld georganiseerd in strakke parallelle bundels door het bundelende eiwit fimbrine. Bundels van de filamenten zijn echter minder dicht opeengepakt, wanneer ze zijn gebonden door alfa-actinine of worden geassocieerd met fibroblast-stressvezels (de parallelle groene vezels in figuur 2). Met name de microfilamentverbindingen die door sommige verknopende eiwitten worden gecreëerd, resulteren in een webachtig netwerk of een gelvorm in plaats van filamentbundels.
In de loop van de evolutionaire geschiedenis van de cel is actine relatief onveranderd gebleven. Dit, samen met het feit dat alle eukaryote cellen sterk afhankelijk zijn van de integriteit van hun actinefilamenten om de vele spanningen waarmee ze in hun omgeving worden geconfronteerd, te kunnen overleven, maakt actine tot een uitstekend doelwit voor organismen die cellen willen beschadigen. Dienovereenkomstig produceren veel planten, die fysiek niet in staat zijn om roofdieren te ontwijken die ze zouden willen opeten of op een andere manier schade zouden willen berokkenen, gifstoffen produceren die cellulaire actine en microfilamenten beïnvloeden als een verdedigingsmechanisme. De doodskap-paddenstoel produceert bijvoorbeeld een stof genaamd phalloidin die zich bindt aan actinefilamenten en deze stabiliseert, wat fataal kan zijn voor cellen.
TERUG NAAR DIER CELSTRUCTUUR
TERUG NAAR DE PLANTENCELSTRUCTUUR
Vragen of opmerkingen? Stuur ons een e-mail.
© 1995-2019 door Michael W. Davidson en The Florida State University. Alle rechten voorbehouden. Afbeeldingen, grafische afbeeldingen, software, scripts of applets mogen op geen enkele manier worden gereproduceerd of gebruikt zonder toestemming van de auteursrechthouders. Gebruik van deze website betekent dat u akkoord gaat met alle juridische voorwaarden die door de eigenaren zijn uiteengezet.
Deze website wordt onderhouden door ons
Graphics & Webprogrammeringsteam
in samenwerking met optische microscopie van het
National High Magnetic Field Laboratory .
Laatste wijziging: vrijdag 13 november 2015 om 13:18 uur
Aantal toegangen sinds 1 oktober 2000: 419769
Microscopen geleverd door:
![]()
![]()