Microfilaments
Communs à toutes les cellules eucaryotes, ces filaments ont une fonction principalement structurelle et sont un composant important du cytosquelette, avec les microtubules et souvent les filaments intermédiaires. Les microfilaments ont un diamètre de 5 à 9 nanomètres et sont conçus pour supporter de grandes quantités de tension. En association avec la myosine, les microfilaments aident à générer les forces utilisées dans la contraction cellulaire et les mouvements cellulaires de base. Les filaments permettent également à une cellule de division de se pincer en deux cellules et sont impliqués dans les mouvements amiboïdes de certains types de cellules.
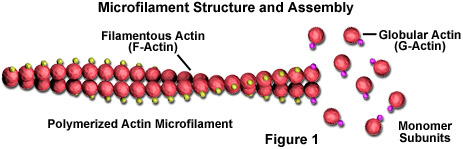
Les microfilaments sont des bâtonnets solides constitués d’une protéine connue sous le nom d’actine. Lorsqu’elle est produite pour la première fois par la cellule, l’actine apparaît sous une forme globulaire (G-actine; voir Figure 1). Dans les microfilaments, cependant, qui sont également souvent appelés filaments d’actine, de longues chaînes polymérisées des molécules sont entrelacées dans une hélice, créant une forme filamenteuse de la protéine (F-actine). Toutes les sous-unités qui composent un microfilament sont connectées de manière à avoir la même orientation. De ce fait, chaque microfilament présente une polarité, les deux extrémités du filament étant nettement différentes. Cette polarité affecte le taux de croissance des microfilaments, une extrémité (appelée l’extrémité plus) s’assemblant et se désassemblant généralement plus rapidement que l’autre (l’extrémité négative).
Contrairement aux microtubules, qui s’étendent généralement à partir du centrosome d’un cellule, les microfilaments sont généralement nucléés au niveau de la membrane plasmique. Par conséquent, la périphérie (bords) d’une cellule contient généralement la plus forte concentration de microfilaments. Cependant, un certain nombre de facteurs externes et un groupe de protéines spéciales influencent les caractéristiques des microfilaments et leur permettent de faire des changements rapides si nécessaire, même si les filaments doivent être complètement démontés dans une région de la cellule et réassemblés ailleurs. Lorsqu’ils se trouvent directement sous la membrane plasmique, les microfilaments sont considérés comme faisant partie du cortex cellulaire, qui régule la forme et le mouvement de la surface de la cellule. Par conséquent, les microfilaments jouent un rôle clé dans le développement de diverses projections de surface cellulaire (comme illustré à la figure 2 ), y compris les filopodes, les lamellipodes et les stéréocils.
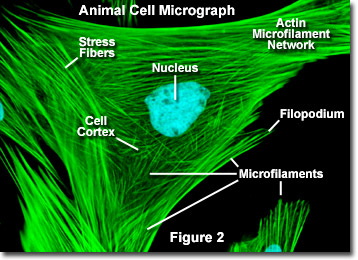
La figure 2 illustre une image numérique par fluorescence d’une cellule de fibroblaste de peau de cerf Muntjac indien colorée avec des sondes fluorescentes ciblant le noyau (bleu) et le réseau cytosquelettique d’actine (vert). Individuellement, les microfilaments sont relativement flexibles. Dans les cellules de organismes vivants, cependant, les filaments d’actine sont généralement organisés en structures plus grandes et beaucoup plus solides par diverses protéines accessoires.La forme structurelle exacte que prend un groupe de microfilaments dépend de leur fonction principale et des protéines particulières qui les lient ensemble. Par exemple, au cœur des saillies de surface appelées microspikes, les microfilaments sont organisés en faisceaux parallèles serrés par la protéine fimbrine. Les faisceaux de filaments sont cependant moins serrés ensemble lorsqu’ils sont liés par l’alpha-actinine ou sont associés à des fibres de tension fibroblastes (les fibres vertes parallèles sur la figure 2). Notamment, les connexions de microfilaments créées par certaines protéines de réticulation aboutissent à un réseau ou une forme de gel de type web plutôt que des faisceaux de filaments.
Au cours de l’histoire évolutive de la cellule, l’actine est restée relativement inchangée. Ceci, ajouté au fait que toutes les cellules eucaryotes dépendent fortement de l’intégrité de leurs filaments d’actine pour pouvoir survivre aux nombreux stress auxquels elles sont confrontées dans leur environnement, fait de l’actine une excellente cible pour les organismes qui cherchent à blesser les cellules. En conséquence, de nombreuses plantes, qui sont incapables d’éviter physiquement les prédateurs qui pourraient vouloir les manger ou leur nuire d’une autre manière, produisent des toxines qui affectent l’actine cellulaire et les microfilaments comme mécanisme défensif. Le champignon mortuaire, par exemple, produit une substance appelée phalloïdine qui se lie aux filaments d’actine et les stabilise, ce qui peut être mortel pour les cellules.
RETOUR À L’ANIMAL STRUCTURE CELLULAIRE
RETOUR À LA STRUCTURE CELLULAIRE VÉGÉTALE
Questions ou commentaires? Envoyez-nous un e-mail.
© 1995-2019 par Michael W. Davidson et la Florida State University. Tous les droits sont réservés. Aucune image, graphique, logiciel, script ou applet ne peut être reproduit ou utilisé de quelque manière que ce soit sans l’autorisation des titulaires des droits d’auteur. L’utilisation de ce site Web signifie que vous acceptez toutes les conditions légales énoncées par les propriétaires.
Ce site Web est géré par notre équipe de programmation Web
Graphics &
en collaboration avec Optical Microscopy au
National High Magnetic Field Laboratory .
Dernière modification: vendredi 13 novembre 2015 à 13:18
Nombre d’accès depuis le 1er octobre 2000: 419769
Microscopes fournis par:
![]()
![]()