Résultats d’apprentissage
- Décrivez comment le potentiel hydrique influence la façon dont l’eau est transportée dans les plantes
Les usines sont des ingénieurs hydrauliques phénoménaux. En utilisant uniquement les lois de base de la physique et la simple manipulation de l’énergie potentielle, les plantes peuvent déplacer l’eau vers le sommet d’un arbre de 116 mètres de haut (figure 1a). Les plantes peuvent également utiliser l’hydraulique pour générer suffisamment de force pour fendre les roches et déformer les trottoirs (figure 1b). Les plantes y parviennent grâce au potentiel hydrique.

Figure 1. Avec des hauteurs proches de 116 mètres, (a ) les séquoias côtiers (Sequoia sempervirens) sont les arbres les plus hauts du monde. Les racines des plantes peuvent facilement générer suffisamment de force pour (b) déformer et briser les trottoirs en béton, au grand désarroi des propriétaires et des services d’entretien de la ville. (crédit a: modification du travail de Bernt Rostad; crédit b: modification du travail de Pedestrians Educating Drivers on Safety, Inc.)
Le potentiel de l’eau est une mesure de l’énergie potentielle dans l’eau. Les physiologistes des plantes ne s’intéressent pas à l’énergie d’un système aqueux en particulier, mais sont très intéressés par le mouvement de l’eau entre deux systèmes. En termes pratiques, le potentiel de l’eau est donc la différence d’énergie potentielle entre un échantillon d’eau donné et de l’eau pure (à la pression atmosphérique et à la température ambiante). Le potentiel de l’eau est désigné par la lettre grecque ψ (psi) et est exprimé en unités de pression (la pression est une forme d’énergie) appelées mégapascals (MPa). Le potentiel de l’eau pure (Ψwpure H2O) est, par commodité de définition, désigné comme une valeur de zéro (même si l’eau pure contient beaucoup d’énergie potentielle, cette énergie est ignorée). Les valeurs du potentiel hydrique de l’eau dans une racine, une tige ou une feuille de plante sont donc exprimées par rapport à Ψwpure H2O.
Le potentiel hydrique des solutions végétales est influencé par la concentration de soluté, la pression, la gravité et des facteurs appelés matrice effets. Le potentiel de l’eau peut être décomposé en ses composants individuels en utilisant l’équation suivante:
Ψsystem = Ψtotal = Ψs + Ψp + Ψg + Ψm
où Ψs, Ψp, Ψg et Ψm se réfèrent respectivement aux potentiels de soluté, de pression, de gravité et de matrice. «Système» peut faire référence au potentiel hydrique de l’eau du sol (Ψsol), de l’eau des racines (Ψracine), de l’eau de la tige (tige), de l’eau des feuilles (Ψleaf) ou de l’eau dans l’atmosphère (Ψatmosphère): quel que soit le système aqueux considéré Lorsque les composants individuels changent, ils augmentent ou diminuent le potentiel d’eau total d’un système. Lorsque cela se produit, l’eau se déplace pour s’équilibrer, passant du système ou du compartiment avec un potentiel d’eau plus élevé au système ou compartiment avec un potentiel d’eau plus faible. Cela ramène la différence de potentiel en eau entre les deux systèmes (ΔΨ) à zéro (ΔΨ = 0). Par conséquent, pour que l’eau se déplace à travers la plante du sol à l’air (un processus appelé transpiration), Ψsoil doit être > Ψroot > Ψstem > Ψleaf > Ψatmosphère .
L’eau se déplace uniquement en réponse à ΔΨ, pas en réponse aux composants individuels. Cependant, parce que les composants individuels influencent le total Ψsystème, en manipulant les composants individuels (en particulier Ψs), une plante peut contrôler le mouvement de l’eau.
Potentiel de soluté
Le potentiel de soluté (Ψs), également appelé potentiel osmotique, est négatif dans un cellule végétale et zéro dans l’eau distillée. Les valeurs typiques du cytoplasme cellulaire sont de –0,5 à –1,0 MPa. Les solutés réduisent le potentiel de l’eau (résultant en un Ψw négatif) en consommant une partie de l’énergie potentielle disponible dans l’eau. Les molécules de soluté peuvent se dissoudre dans l’eau parce que les molécules d’eau peuvent s’y lier via des liaisons hydrogène; une molécule hydrophobe comme l’huile, qui ne peut pas se lier à l’eau, ne peut pas se dissoudre. L’énergie contenue dans les liaisons hydrogène entre les molécules de soluté et l’eau n’est plus disponible pour travailler dans le système car elle est liée à la liaison. En d’autres termes, la quantité d’énergie potentielle disponible est réduite lorsque des solutés sont ajoutés à un système aqueux. Ainsi, Ψs diminue avec l’augmentation de la concentration en soluté. Puisque Ψs est l’un des quatre composants de Ψsystem ou Ψtotal, une diminution de Ψs entraînera une diminution de Ψtotal. Le potentiel hydrique interne d’une cellule végétale est plus négatif que l’eau pure en raison de la teneur élevée en soluté du cytoplasme (figure 2). En raison de cette différence de potentiel hydrique, l’eau se déplacera du sol vers les cellules racinaires d’une plante via le processus d’osmose. C’est pourquoi le potentiel de soluté est parfois appelé potentiel osmotique.
Les cellules végétales peuvent manipuler métaboliquement Ψs (et par extension, Ψtotal) en ajoutant ou en supprimant des molécules de soluté. Par conséquent, les plantes contrôlent overtotal via leur capacité à exercer un contrôle métabolique sur Ψs.
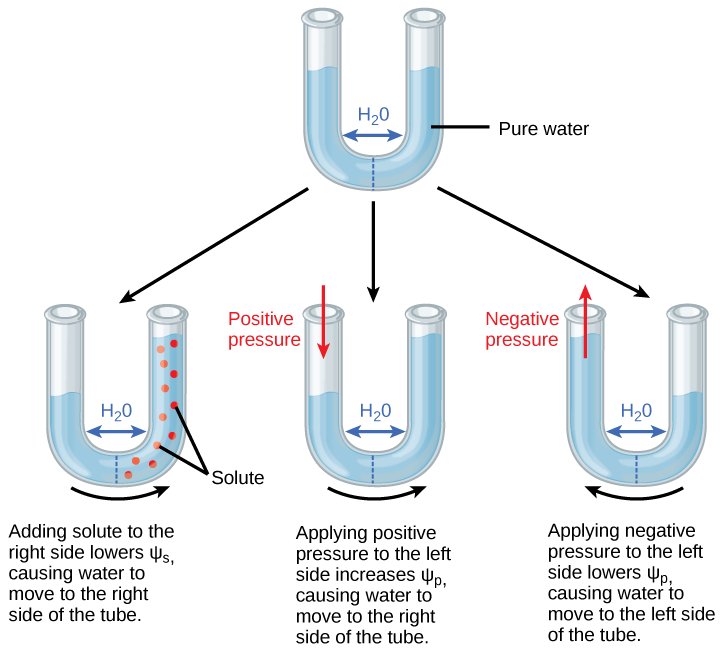
Figure 2 .Une membrane semi-perméable entre deux systèmes aqueux
Dans la figure 2, l’eau se déplacera d’une région de potentiel hydrique supérieur à inférieur jusqu’à ce que l’équilibre soit atteint. Les solutés (Ψs), la pression (Ψp) et la gravité (Ψg) influencent le potentiel d’eau total de chaque côté du tube (Ψ total à droite ou à gauche), et par conséquent, la différence entre Ψtotal de chaque côté (ΔΨ). (Ψm, le potentiel dû à l’interaction de l’eau avec des substrats solides, est ignoré dans cet exemple car le verre n’est pas spécialement hydrophile). L’eau se déplace en réponse à la différence de potentiel d’eau entre deux systèmes (les côtés gauche et droit du tube).
Question pratique
Le potentiel d’eau positif est placé sur le côté gauche du tube en augmentant Ψp de telle sorte que le niveau d’eau monte du côté droit. Pourriez-vous égaliser le niveau d’eau de chaque côté du tube en ajoutant du soluté, et si oui, comment?
Potentiel de pression
Le potentiel de pression (Ψp), également appelé potentiel de turgescence, peut être positif ou négatif (Figure 2). Parce que la pression est une expression d’énergie, plus la pression est élevée, plus il y a d’énergie potentielle dans un système, et vice versa. Par conséquent, un Ψp positif (compression) augmente Ψtotal, et un Ψp (tension) négatif diminue Ψtotal. La pression positive à l’intérieur des cellules est contenue par la paroi cellulaire, produisant une pression de turgescence. Les potentiels de pression sont généralement de l’ordre de 0,6 à 0,8 MPa, mais peuvent atteindre 1,5 MPa dans une plante bien arrosée. Un Ψp de 1,5 MPa équivaut à 210 livres par pouce carré (1,5 MPa x 140 lb in-2 MPa-1 = 210 lb / in-2). À titre de comparaison, la plupart des pneus automobiles sont maintenus à une pression de 30 à 34 psi. Un exemple de l’effet de la pression de turgescence est le flétrissement des feuilles et leur restauration après que la plante a été arrosée (Figure 3). L’eau est perdue des feuilles par transpiration (approchant Ψp = 0 MPa au point de flétrissement) et restituée par absorption via les racines.
Une plante peut manipuler Ψp via sa capacité à manipuler Ψs et par le processus de osmose. Si une cellule végétale augmente la concentration de soluté cytoplasmique, Ψs diminuera, Ψtotal diminuera, le ΔΨ entre la cellule et le tissu environnant diminuera, l’eau se déplacera dans la cellule par osmose et Ψp augmentera. Ψp est également sous contrôle indirect des plantes via l’ouverture et la fermeture des stomates. Les ouvertures stomatiques permettent à l’eau de s’évaporer de la feuille, réduisant Ψp et Ψtotal de la feuille et augmentant ii entre l’eau de la feuille et le pétiole, permettant ainsi à l’eau de s’écouler du pétiole dans la feuille.

Figure 3. Lorsque (a) le potentiel d’eau total (Ψtotal) est plus faible à l’extérieur des cellules qu’à l’intérieur, l’eau sort des cellules et la plante se flétrit. Lorsque (b) le potentiel d’eau total est plus élevé à l’extérieur des cellules végétales qu’à l’intérieur, l’eau pénètre dans les cellules, ce qui entraîne une pression de turgescence (Ψp) et maintient la plante debout. (crédit: modification de l’oeuvre de Victor M. Vicente Selvas)
Potentiel de gravité
Le potentiel de gravité (Ψg) est toujours négatif à zéro dans une plante sans hauteur. Il supprime ou consomme toujours de l’énergie potentielle du système. La force de gravité tire l’eau vers le sol, réduisant la quantité totale d’énergie potentielle dans l’eau de la plante (Ψtotal). Plus la plante est haute, plus la colonne d’eau est haute et plus Ψg devient influent. A l’échelle cellulaire et chez les plantes courtes, cet effet est négligeable et facilement ignoré. Cependant, sur la hauteur d’un grand arbre comme un séquoia géant côtier, l’attraction gravitationnelle de –0,1 MPa m-1 équivaut à 1 MPa supplémentaire de résistance qui doit être surmontée pour que l’eau atteigne les feuilles des arbres les plus hauts. Les plantes sont incapables de manipuler Ψg.
Potentiel matriciel
Le potentiel matriciel (Ψm) est toujours négatif à zéro. Dans un système sec, il peut être aussi bas que –2 MPa dans une graine sèche, et il est nul dans un système saturé d’eau. La liaison de l’eau à une matrice supprime ou consomme toujours de l’énergie potentielle du système. Ψm est similaire au potentiel de soluté car il implique de lier l’énergie dans un système aqueux en formant des liaisons hydrogène entre l’eau et un autre composant. Cependant, en potentiel de soluté, les autres composants sont des molécules de soluté hydrophiles solubles, alors qu’en Ψm, les autres composants sont des molécules hydrophiles insolubles de la paroi cellulaire végétale. Chaque cellule végétale a une paroi cellulaire cellulosique et la cellulose dans les parois cellulaires est hydrophile, produisant une matrice d’adhésion de l’eau: d’où le nom potentiel matriciel. Ψm est très grand (négatif) dans les tissus secs tels que les graines ou les sols affectés par la sécheresse. Cependant, il passe rapidement à zéro lorsque la graine absorbe de l’eau ou que le sol s’hydrate.Ψm ne peut pas être manipulé par la plante et est généralement ignoré dans les racines, tiges et feuilles bien arrosées.
Essayez
Contribuez!
Améliorez cette pageEn savoir plus