Microfilamentos
Comum a todas as células eucarióticas, esses filamentos têm função principalmente estrutural e são um componente importante do citoesqueleto, junto com os microtúbulos e, frequentemente, os filamentos intermediários. Os microfilamentos variam de 5 a 9 nanômetros de diâmetro e são projetados para suportar grandes quantidades de tensão. Em associação com a miosina, os microfilamentos ajudam a gerar as forças usadas na contração celular e nos movimentos celulares básicos. Os filamentos também permitem que uma célula em divisão se separe em duas células e estão envolvidos nos movimentos amebóides de certos tipos de células.
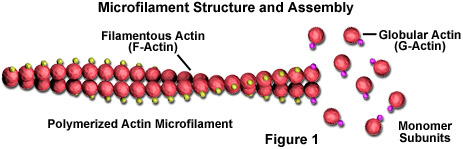
Microfilamentos são bastonetes sólidos feitos de uma proteína conhecida como actina. Quando é produzida pela primeira vez pela célula, a actina aparece na forma globular (G-actina; ver Figura 1). Em microfilamentos, entretanto, que também são freqüentemente referidos como filamentos de actina, longas cadeias polimerizadas das moléculas são entrelaçadas em uma hélice, criando uma forma filamentosa da proteína (F-actina). Todas as subunidades que compõem um microfilamento estão conectadas de forma que tenham a mesma orientação. Devido a este fato, cada microfilamento exibe polaridade, as duas extremidades do filamento sendo distintamente diferentes. Esta polaridade afeta a taxa de crescimento dos microfilamentos, uma extremidade (denominada extremidade positiva) normalmente se monta e desmonta mais rápido do que a outra (extremidade negativa).
Ao contrário dos microtúbulos, que normalmente se estendem a partir do centrossoma de um célula, os microfilamentos são tipicamente nucleados na membrana plasmática. Portanto, a periferia (bordas) de uma célula geralmente contém a maior concentração de microfilamentos. Uma série de fatores externos e um grupo de proteínas especiais influenciam as características do microfilamento, no entanto, e permitem que eles façam mudanças rápidas se necessário, mesmo se os filamentos precisarem ser completamente desmontados em uma região da célula e remontados em outro lugar. Quando encontrados diretamente abaixo da membrana plasmática, os microfilamentos são considerados parte do córtex celular, que regula a forma e o movimento da superfície celular. Consequentemente, os microfilamentos desempenham um papel fundamental no desenvolvimento de várias projeções da superfície celular (conforme ilustrado na Figura 2 ), incluindo filopódios, lamelipódios e estereocílios.
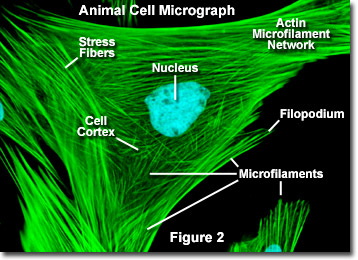
Ilustrado na Figura 2 está uma imagem digital de fluorescência de uma célula de fibroblasto de pele de veado indiano Muntjac corada com sondas fluorescentes direcionadas ao núcleo (azul) e à rede do citoesqueleto de actina (verde). Individualmente, os microfilamentos são relativamente flexíveis. Nas células de organismos vivos, no entanto, os filamentos de actina são geralmente organizados em estruturas maiores e muito mais fortes por várias proteínas acessórias. A forma estrutural exata que um grupo de microfilamentos assume depende de sua função primária e das proteínas específicas que os unem. Por exemplo, no núcleo das protrusões superficiais chamadas micro-pontas, os microfilamentos são organizados em feixes paralelos pela proteína fimbrina. Feixes de filamentos são menos compactados, no entanto, quando são ligados por alfa-actinina ou associados a fibras de estresse de fibroblastos (as fibras verdes paralelas na Figura 2). Notavelmente, as conexões de microfilamento criadas por algumas proteínas de reticulação resultam em uma rede semelhante a uma teia ou forma de gel em vez de feixes de filamentos.
Ao longo da história evolutiva da célula, a actina permaneceu relativamente inalterada. Isso, junto com o fato de que todas as células eucarióticas dependem fortemente da integridade de seus filamentos de actina para poder sobreviver aos muitos estresses que enfrentam em seu ambiente, torna a actina um excelente alvo para organismos que procuram ferir células. Conseqüentemente, muitas plantas, incapazes de evitar fisicamente os predadores que possam querer comê-las ou prejudicá-las de alguma outra forma, produzem toxinas que afetam a actina celular e os microfilamentos como mecanismo de defesa. O cogumelo da tampa da morte, por exemplo, produz uma substância chamada faloidina que se liga e estabiliza os filamentos de actina, o que pode ser fatal para as células.
VOLTAR AO ANIMAL ESTRUTURA CELULAR
VOLTAR À ESTRUTURA CELULAR
Perguntas ou comentários? Envie-nos um e-mail.
© 1995-2019 por Michael W. Davidson e The Florida State University. Todos os direitos reservados. Nenhuma imagem, gráfico, software, script ou miniaplicativo pode ser reproduzido ou usado de qualquer maneira sem a permissão dos detentores dos direitos autorais. O uso deste site significa que você concorda com todos os Termos e Condições Legais estabelecidos pelos proprietários.
Este site é mantido por nossa
Graphics & Equipe de programação da Web
em colaboração com Microscopia Óptica no
National High Magnetic Field Laboratory .
Última modificação: sexta-feira, 13 de novembro de 2015 às 13h18
Contagem de acessos desde 1º de outubro de 2000: 419769
Microscópios fornecidos por:
![]()
![]()